



Rhysodini
Wrinkled Bark Beetles
Ross T. Bell


Introduction
Rhysodini is a highly specialized carabid tribe formerly ranked as a separate family, Rhysodidae. It contains about 350 known species in seven subtribes. It occurs nearly world wide in suitably moist tropical and temperate forests.
These beetles are difficult to collect in numbers. They are most often encountered under loose bark of dead trees, in stumps and fallen logs, also in the sawdust-like debris within dead hollow trees. This has given a misleading impression of the usual habitat. The beetles actually live between the layers of dead wood where they can be collected by splitting the wood. They do not make burrows but push themselves between the layers, compressing the wood cells as they go. The cells rebound after they pass, leaving no obvious trace of their passage. The exoskeleton is very thick and strong, resisting the pressure of the wood. The beetles are thought to live on the amoeboid stages of slime molds (Myxomycetes).
More than half of the rhysodine species have fully developed hind wings, but they have rarely been caught at light traps, and then only within forests. Probably flight is used primarily for reaching dead areas in the upper parts of trees, rather than for dispersal. On rare occasions, adults of vestigial-winged species have been caught in pitfall traps in forests, on the surfaces of fallen logs at night, or in forest litter. In addition to dead trees, they have been taken in dead parts of living trees, including branches which have just fallen from the canopy. They are also found occasionally in roots, sometimes as deep as three meters. They usually occur in groups; if one is found, it can be profitable to dissect as much of the log or stump as is practical.
All species vary from dark red to nearly black; color appears to deepen with age. In some species, the eyes are clear and seemingly functional in the lighter, presumably younger specimens, but become obscured by dark pigment in the darker specimens.
The larvae live in short tunnels within dead wood. The tunnel behind the larva becomes packed with wood fragments. Larvae are found only infrequently. They are commonly in the same wood that contains adults. Diversity of larvae is almost unstudied.
Characteristics
Adults
The body is long and narrow, with a relatively long prothorax. The antennae are short for Adephaga, reaching only halfway to the base of the pronotum, but they are thick and relatively stiff. The segments are moniliform (shaped like beads). The head has a large internal cavity which opens to the exterior by a pore in the occipital region or by various arrangements of fissures which meet in the occipital region. In dorsal view, the head usually consists of a median lobe flanked by a pair of temporal lobes, each bearing a compound eye. The base of the head forms a large cervical condyle.
The mouthparts are unique. The labrum is triangular, with four or two setae. The mandibles are thin, and without cutting edges. They serve as covers for the other mouthparts but are not used in feeding. Each mandible has a scrobal seta and a hollowed external tooth, the auricle. When the mandibles are closed, the auricles are nested above the large median tooth of the mentum. The mentum is fused laterally to the head capsule. Anteriorly, it projects beyond the other mouthparts with its anterior margin forming the cutting edge as the beetle forces its way through the wood. The labial palpi are minute. They are hidden by the mentum when the jaws are closed but emerge when the jaws are open. The maxillae and their palpi are also completely hidden when the jaws are shut, except for the tip of the palpus. The galea and lacinia of the maxillae are converted into stylets.
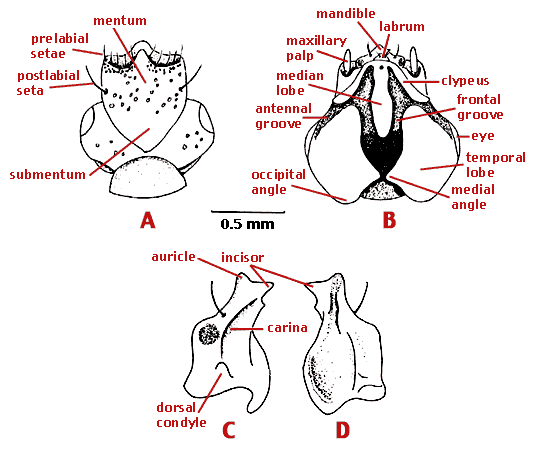
Mouthparts of rhysodines. A. Head of Omoglymmius sp., ventral view. B. Dorsal view. C. Left mandible of Clinidium sp., ventral view. D. dorsal view.
Modified from Bell (1994). Copyright © 1994 Ross T. Bell.
The eyes are extremely variable in form. They are large in fully winged species, but, unlike those of other Carabidae, they do not project from the head. They may be very small in flightless species but are never entirely absent.
The pronotum is elongate. The median groove and basal impressions are usually deeply incised; in many species, the basal impressions are extended to the anterior margin, dividing the surface of the pronotum into four raised ridges, the inner and outer carinae. The pronotum only rarely has a sharp lateral edge, but it usually has a marginal groove, or two such grooves, delineating a narrow marginal carina. The pronotal sutures are distinct, but the prosternal suture is absent, so the prosternum and propleura are fused. Often there is a groove in place of the prosternal suture, but it is not a true suture. The mesothorax is narrowed and elongate as in Scaritini. Each elytron has eight or fewer striae. The scutellar striole is absent. The meso- and metacoxal cavities are disjunct. The abdomen appears to have five visible sterna. Comparison with the dorsal surface of the abdomen reveals that the apparent second sternum actually represents a fusion of the second and third sterna.
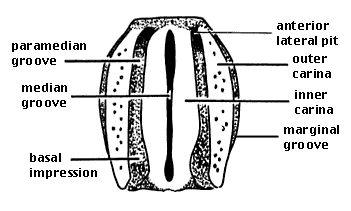
Dorsal aspect of a rhysodine pronotum (diagrammatic).
Modified from Bell and Bell (1978). Copyright © 1978 Ross T. Bell and Joyce R. Bell.
The hind coxae are relatively small and are further apart than in most Carabidae, so the median remnant of Sternum I is larger than usual. The metasternum lacks a transverse suture, and the median groove varies from a moderate sulcus to non-existent.
The legs are relatively short but are very strongly built. The front tibia has two medially curved apical processes (anterior and posterior). A well-developed cleaning organ is present. It is of an advanced type, more distal than in other Carabidae, and its base varies from opposite the tarsal insertion to entirely distal to it. The cleaning organ is a curved row of hairs without clip setae or obvious tibial spurs. (There is a minute tibial spur at the proximal end of the cleaner in a few species, demonstrating derivation from the anisochaetous type of cleaning organ.) The outer distal angle of the middle tibia is acute or else it forms a medially curved process. The male front tarsi are not modified. The middle and hind tibiae of the male each have a calcar, a distal, anteriorly directed process. This character provides a completely reliable method of determining the sex of specimens.
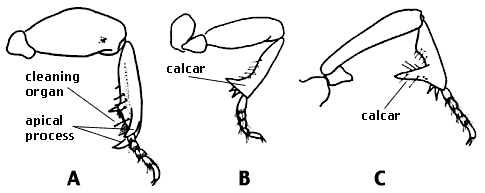
Fore (A), middle (B), and hind leg (C) of a Clinidium calcaratum male.
Modified from Bell (1970). Copyright © 1970 Entomological Society of America.
The male genitalia have highly unusual parameres which lack setae (similar to the Harpalinae). The form of the penis and the development of the internal sac are highly varied. The tactile setae are well developed. They are similar in arrangements to other carabid groups except that the specialized margin (umbilical) series is absent. Parts of the surface of the exoskeleton are covered with pollinosity (extremely minute scales or hairs to which some wood fibers are usually adherent).
Larvae
The larva is pale, soft-bodied, and 9 mm or less in length. It is grublike without distinct sclerites on thorax or abdomen and without urogomphi. Legs are short but have six segments, as is usual in Adephaga. There is only one tarsal claw. Most of the body segments each have a dorsal transverse row of spinulae which is interrupted at the midline. At a minimum there are such rows on the metathorax and abdominal segments 1-6. Eyes are absent. There is a soft epipharynx projecting anteriorly. In the past, this has been mistaken for a labrum. The nasale is truncate, the labial palpi are reduced to vestiges, and the maxillae and prementum bear fringed membranes.
Relationships of Rhysodini to Other Beetles
These highly modified beetles have been moved from place to place among the Coleoptera. At first they were grouped with other wood-dwelling beetles of somewhat similar appearance, and therefore, among the Polyphaga. The wing venation, the larval leg segmentation, and other characters demonstrated that they belong among Adephaga. Jeannel and others considered that they should form an independent, and presumably primitive superfamily within the suborder. Bell and Bell (1962) doubted this and placed them as a tribe of Carabidae with some sort of relationship to Scaritini. Recently, Bell (1998) produced evidence that they belong within Scaritinae, in the broad sense, and that the sister group is the Neotropical genus Solenogenys, usually placed in the subtribe Salcediina (placed in Clivinini in the Tree of Life project).
Discussion of Phylogenetic Relationships
Interrelationships among the seven subtribes of Rhysodini remain to be worked out. Potential sister groups of each of the subtribes are discussed on the subtribal pages.
Geographic Distribution
Rhysodini occur on all continents and most islands that have suitable forests. Outer limits are New Zealand, Fiji, and Kusiae (in the Eastern Carolines). In North America they can be found north to Vancouver Island, Illinois, and the south shore of Lake Ontario. They are present throughout the West Indies. In Eurasia, Rhysodini occur north to southern Sweden (perhaps now extinct there), Poland, western China, and southeastern Siberia. They are also found in South America, south to Ecuador and Paraguay (but they are unaccountably absent from the temperate forests of Chile). They are present in the forested parts of Africa, Madagascar, and Ceylon, as well as all parts of the Philippines and Indonesia. In Australia, they are found on the East Coast from Cape York to Tasmania. The fauna is richest in species in New Guinea, Indonesia, the Philippines, and northern South America. Southeast Asia and Indonesia are most diverse in genera.
References
Bell, R. T. 1970. The Rhysodini of North America, Central America and the West Indies. Miscellaneous Publications of the Entomological Society of America 6: 289-324.
Bell, R. T. 1973. New species of Clinidium from Guatemala (Coleoptera, Carabidae or Rhysodidae). Proceedings of the Entomological Society of Washington 75(3): 279-282.
Bell, R. T. 1975. Omoglymmius Ganglbauer, a separate genus (Coleoptera: Carabidae or Rhysodidae). The Coleopterists Bulletin 29(4):351-352.
Bell, R. T. 1979. Zoogeography of Rhysodini: Do beetles travel on driftwood? Pages 331-342 in Carabid Beetles: Their Evolution, Natural History and Classification (T. L. Erwin, G. E. Ball, and D. R. Whitehead, eds.). Proceedings of the First International Symposium of Carabidology. Smithsonoan Institution, Wahsington, D.C.
Bell, R. T. 1985. Zoogeography and ecology of New Guinea Rhysodini (Coleoptera: Carabidae). Pages 221-235 in Taxonomy, Phylogeny and Zoogeography of Beetles and Ants (G. E. Ball, ed.). Dr. W. Junk Publishers, Dordrecht.
Bell, R. T. 1985. Family: Rhysodidae. A Catalog of the Coleoptera of America North of Mexico. United States Department of Agriculture. Agriculture Handbook Number 529-4:1-4.
Bell, R. T. 1991. Rhysodidae (Adephaga). Pages 304-305 in Immature Insects Volume 2 (F. W. Stehr, ed.). Kendall/Hunt Publishing Company, Dubuque, Iowa.
Bell, R. T. 1994. Beetles that cannot bite: functional morphology of the head of adult Rhysodines (Coleoptera: Carabidae or Rhysodidae). The Canadian Entomologist 126:667-672.
Bell, R. T. 1998. Where do the Rhysodini (Coleoptera) belong? Pages 261-272 in Phylogeny and Classification of Caraboidea. XX I.C.E. (1996, Firenze, Italy) (G. E. Ball, A. Casale, and A. Vigna Taglianti, eds.). Museo Regionale di Scienze Naturali, Torino.
Bell, R. T. and J. R. Bell. 1962. The taxonomic position of the Rhysodidae. The Coleopterists Bulletin 15:99-106.
Bell, R. T. and J. R. Bell. 1975. Two new taxa of Clinidium (Coleoptera: Rhysodidae or Carabidae) from the Eastern U.S., with a revised key to U.S. Clinidium. The Coleopterists Bulletin 29(2):65-68.
Bell, R. T. and J. R. Bell. 1978. Rhysodini of the World part I. A new classification of the tribe, and a synopsis of Omoglymmius subgenus Nitiglymmius, new subgenus (Coleoptera: Carabidae or Rhysodidae). Quaestiones Entomologicae 14: 43-88.
Bell, R. T. and J. R. Bell. 1979. Rhysodini of the World part II. Revisions of the smaller genera (Coleoptera: Carabidae or Rhysodidae). Quaestiones Entomologicae 15: 377-446.
Bell, R. T. and J. R. Bell. 1982. Rhysodini of the World Part III. Revision of Omoglymmius Ganglbauer (Coleoptera: Carabidae or Rhysodidae) and substitutions for preoccupied generic names. Quaestiones Entomologicae 18: 127-259.
Bell, R. T. and J. R. Bell. 1985. Rhysodini of the World Part IV. Revisions of Rhyzodiastes and Clinidium, with new species in other genera (Coleoptera: Carabidae or Rhysodidae). Quaestiones Entomologicae 21(1): 1-172.
Bell, R. T. and J. R. Bell. 1987. A new species of Clinidium Kirby (Coleoptera: Carabidae or Rhysodidae) from Mexico, and descriptions of the females of two Neotropical members of the genus. Annals Of The Carnegie Museum 56(8-14): 193-196.
Bell, R. T. and J. R. Bell. 1987. A new subtribe, genus and species of Rhysodini from South Africa (Coleoptera: Carabidae or Rhysodidae). Journal Of The Entomological Society Of Southern Africa 50(2): 287-290.
Bell, R. T. and J. R. Bell. 1987. Rhysodine beetles on the Geneva collection: a new species of Yamatosa, and a major range extension for Omoglymmius sakuraii Nakane (Coleoptera: Carabidae or Rhysodidae). Revue suisse Zool. 94(4):683-686.
Bell, R. T. and J. R. Bell. 1988. Rhysodini of Sulawesi (Indonesia) and nearby islands (Coleoptera: Carabidae or Rhysodidae). Journal Of The New York Entomological Society 96(1): 7-15.
Bell, R. T. and J. R. Bell. 1989. Rhysodine beetles on the Geneva collection II: new species of Yamatosa and Omoglymmius, descriptions of undescribed sexes in other species, and some major range extensions (Coleoptera: Carabidae or Rhysodidae). Revue suisse Zool. 96(3):637-642.
Bell, R. T. and J. R. Bell. 1991. The Rhysodini of Australia (Insecta: Coleoptera: Carabidae or Rhysodidae). Annals Of The Carnegie Museum 60(3): 179-210.
Bell, R. T. and J. R. Bell. 1993. Rhysodine beetles (Insecta: Coleoptera: Carabidae or Rhysodidae): New species, new data, and revised keys to Omoglymmius (Subgenera Omoglymmius and Pyxiglimmius). Annals of Carnegie Museum 62(2): 165-185 .
Bell, R. T. and J. R. Bell. 1995. The Rhysodini (Insecta: Coleoptera: Carabidae) of Cuba. Annals of Carnegie Museum 64(3): 185-195.
Beutel, R. G. 1990. Metathoracic features of Omoglymmius hamatus and their significance for classification of Rhysodini (Coleoptera: Adephaga). Entomologia Generalis 15(3): 185-201.
Beutel, R. G. 1992. Larval head structures of Omoglymmius hamatus and their implications for the relationships of Rhysodidae (Coleoptera: Adephaga). Entomologica Scandinavica 23(2): 169-184.
Bousquet, Y. and A. Larochelle. 1993. Catalogue of the Geadephaga (Coleoptera: Trachypachidae, Rhysodidae, Carabidae including Cicindelini) of America north of Mexico. Memoirs of the Entomological Society of Canada 0(167):3-334.
Burakowski, B. 1975. Description of larva and pupa of Rhysodes sulcatus (F.) (Coleoptera, Rhysodidae) and notes on the bionomics of this species. Annales Zoologici 32(12)271-287.
Costa, C., S. A. Vanin, and S. A. Casari-Chen. 1988. Larvas de Coleoptera do Brasil. 269 pp. Sao Paolo.
Emberson, R. M. 1995. A new species of Kaveinga (Coleoptera: Rhysodidae) and lectotype designations for four species of rhysodids described by Thomas Broun (1880). New Zealand Entomologist 18(0):9-14.
Grandi, G. 1972. Comparative morphology and ethology of insects with a specialized diet, Rhysodes germari Ganglb. Bolletino dell'Istituto di Entomologia dell' Università di Bologna 30:21-47.
Hovorka, 0. 1997. New Clinidium species from Ecuador (Coleoptera: Carabidae: Rhysodini). Acta Societatis Zoologicae Bohemicae 61(1):19-22.
Yahiro, K. 1996. Comparative morphology of the alimentary canal and reproductive organs of the terrestrial Caraboidea (Coleoptera: Adephaga) Part 1. Japanese Journal of Entomology 64(3):536-550.
Title Illustrations

| Scientific Name | Omoglymmius hamatus |
|---|---|
| Location | California, U.S.A. |
| Specimen Condition | Live Specimen |
| Image Use |
 This media file is licensed under the Creative Commons Attribution License - Version 3.0. This media file is licensed under the Creative Commons Attribution License - Version 3.0.
|
| Copyright |
© 2000 David R. Maddison

|

| Scientific Name | Clinidium baldufi |
|---|---|
| Location | Mississippi, U.S.A. |
| Specimen Condition | Live Specimen |
| Image Use |
This media file is licensed under the Creative Commons Attribution License - Version 3.0.
|
| Copyright |
© 1997 David R. Maddison
|
About This Page
Ross T. Bell
University of Vermont, Burlington, Vermont, USA
Page copyright © 1999 Ross T. Bell
All Rights Reserved.
- First online 14 December 1999
Citing this page:
Bell, Ross T. 1999. Rhysodini. Wrinkled Bark Beetles. Version 14 December 1999. http://tolweb.org/Rhysodini/67/1999.12.14 in The Tree of Life Web Project, http://tolweb.org/